
Welfare Biology and AI: What We Can Do Now
Chapter 3: From soil-welfare research to the New World Screwworm: a practical portfolio for wild-invertebrate welfare.


This is part 3 of a five-part sequence on welfare ecology. Part 1 introduces the ethical premises. Part 2 covers the empirical landscape. Part 4 explores a model of invertebrate suffering. Part 5 covers AI.
In part 2, I argued that land use is a key lever for wild-invertebrate welfare and that pesticides applied at constant net primary productivity (NPP) can create a high-throughput killing field rather than a real population reduction. This post turns to the practical question: given that picture, what should we do?
The answer depends on which premises from part 1 you hold. I’ll write from my own view – asymmetric antifrustrationism with non-negligible probability mass on invertebrate sentience – but I’ll flag where the conclusions change for symmetric utilitarians or for people who don’t extend moral concern below the arthropod line.
A month ago, Rethink Priorities (RP) published its Wild Animal Welfare Intervention Database (WAWID), twenty-eight shallow reports on candidate interventions. It changes the picture. Several of the most promising entries are birth-prevention interventions that map cleanly onto the framework from parts 1 and 2, and a couple are surprises – the New World Screwworm (NWS) suppression case in particular is, on my read, the single strongest entry in the database.
A Key Lever: Land Use
Part 2 argued that the m²-years of agricultural land per food-kg explain almost all of the variance in welfare effects across foods. Converting natural biomes to agriculture reduces soil fauna density by 2–8×, and the sheer number of organisms affected (billions per m²-year) overwhelms everything else.
That mechanism is what makes land use change – not pesticide use, not “humane mowing,” not habitat enrichment – the central lever. Most of the interventions discussed below either work directly through it (existing donations, dietary choices, biofuels as a side effect) or are picked specifically because they do something land use cannot, like target a fly that causes excruciating pain to mammals it parasitizes.
Two Asymmetries to Filter By
Before we go through the candidates, two filters from the asymmetric antifrustrationist position do most of the prioritizing work. They aren’t decorative – they sharply rerank what looks promising.
Pro-choice. Or birth-prevention beats death-improvement: Under the asymmetry from part 1, creating frustrated lives is bad; preventing them isn’t. Improving an existing being’s mode of death matters, but the welfare delta is bounded by the difference between two deaths – a few hours of pyrethroid neurotoxicity versus a few hours of fungal infection, say. This has advantages for collaborations across ethical camps, though. Preventing that being from existing in the first place is bounded by their entire potential life. For r-strategists with high fecundity and short life expectancy at birth (LEB), almost the entire LEB is pre-reproductive death anyway, so the “future-life” side of the trade is large.
Soft Pareto. Or creating new frustrated lives in service of preventing others is bad: Bounded marginal costs to existing beings – slightly more expensive renewable fuel, a small Pigouvian tax, soil disturbance during a one-time land-conversion event whose victims would have died of other causes within weeks – are the kinds of trades society routinely makes. But bringing new sentient beings into existence as the means by which welfare gains for others are achieved is a different kind of action. The asymmetry weighs the entire frustrated life that gets created against the prevented frustrations elsewhere; there’s no bounded “marginal” framing that contains the harm. This is why eating beef to displace soy production registers differently from biofuel subsidies that reduce soil-fauna populations as a policy side effect: the first creates cattle in order to prevent invertebrate births; the second imposes bounded human costs (slightly higher food prices) to produce the same kind of welfare benefit.
There are two important caveats to this:
The bearability buffer. I consider a slight increase in taxes to be a small cost and suffering to be a great cost, largely because my intuiton is that macroscopic welfare ranges abound and that everyone has a kind of individual bearability buffer where costs remain costs and don’t cross over into suffering unless they become chronic. People who go along with the dust speck argument might not feel like there is such a bearability buffer.
The counterfactuals matter. If I have some money, and I want to decide whether to donate it to the Shrimp Welfare Project or the Aquatic Life Institute, I can make a decision, and I’ll impose an easily bearable cost on myself to save some shrimps or fish. But say I decide to save some fish, but I think about it for another day and then redecide and now rather want to save some shrimp. Suddenly I’m sacrificing fish for shrimp. The bearable cost to myself has become an unbearable cost to the fish only because I’ve changed the counterfactual.
Taken together, these considerations confuse me. The first is clear enough, but once the second enters the picture I reliably lose my ethical footing. For the time being, I think of it along the lines of contractualism. If I can reasonably expect that someone expects me to make a certain choice, then that choice is my default to compare others against. In cases where there is no such expectation, I at least don’t judge people for the counterfactuals they choose.
As an example, humans in need sort of vaguely expect to perhaps be helped. Humans in general sort of vaguely expect not to be killed. But serial donors and serial killers are a thing. When a serial donor decides to donate somewhere, the decide not to donate elsewhere; likewise, when a serial killer decides to spare someone they decide to kill someone else. Naturally, in the age of financial self-determination, mood stabilizers, antipsychotics, and Dialectical Behavioral Therapy, we’re not locked into our nature, be it that of the serial donor or the serial killer – any serial killer could become a serial donor if they put their mind to it – but bracketing for the moment these changes to the whole setup, they face very similar decision: who gets to live and who gets to die – only serial donors get to make it at a vastly greater scale. This consideration has made me a lot less judgmental of serial killers.
In summary, the two filters – pro-choice and soft Pareto – favor: birth-prevention interventions, broad-spectrum population-reducing levers, direction-agnostic research, and bounded-marginal-cost trades. They disfavor: interventions that only mildly improve existing deaths without reducing future births, conservation-style interventions that increase aggregate populations, and any structure where new sentient beings are brought into existence to improve the welfare arithmetic.
A third filter follows from part 2’s empirical picture: prefer interventions that affect both nematodes and arthropods, because the dominant uncertainty in any cost-effectiveness analysis here is the welfare range of nematodes – the most numerous probably-sentient animals on Earth, and the ones we know least about. An intervention that is good whether nematodes turn out to have substantial or negligible welfare ranges is more robust than one whose whole raison d’être rests on that question.
Foundational Research
Research stikes me as the most cost-effective category, and that’s why it goes first. The reason is structural: the entire portfolio below depends on comparisons of welfare ranges and on how fine-grained land-use changes propagate through trophic networks. A few hundred thousand dollars of well-targeted welfare-biology research could redirect tens or hundreds of millions of dollars of downstream allocation.
Three concrete research directions look strongest.
Soil-animal welfare research. This is the question the rest of the field is logically downstream of. We need functional characterization of nociception in nematodes – the connectome of Caenorhabditis elegans is mapped, but we don’t know whether its nociceptive processing produces phenomenal pain – along with welfare-per-animal-year estimates for soil mites and springtails, and on-the-ground measurements of how specific land-use changes propagate to soil-fauna densities and whether they are stable or wash out over time (short of millions of years) or get offset in some way. The current cost-effectiveness numbers, including the welfare range of 6.68 × 10⁻⁶ for nematodes that Grilo deferred to Gemini 2.5 to estimate, are placeholders waiting for empirical replacement. The Wild Animal Initiative (WAI) and Rethink Priorities are the obvious organizations to defer to.
Insecticide welfare research. The WAWID’s research-agenda entry by Simon Eckerström Liedholm proposes comparing insecticide classes on welfare effects on wild insects, not just mortality. Roughly 3.5 × 10¹⁵ insects are exposed to insecticides on US farmland each year. We have no idea which classes minimize per-individual suffering – pyrethroids cause hours of paralytic seizure, neonicotinoids 1–2 days of disorientation and twitching, Bacillus thuringiensis (Bt) leads to multi-day death by sepsis, insect growth regulators kill at the next molt by failure to ecdyse. Picking the least-bad class is direction-agnostic: the recommendation holds whether wild insect lives are net negative (then we want euthanasia, but as gentle as possible) or net positive (then we ought not to interfere, but failing that have a duty to minimize the suffering we impose).
Field-building. Mal Graham’s WAWID shallow on scientific field-building for wild animal welfare frames a different version of the research question: what does it cost to bring the field to maturity, with degree programs, peer-reviewed journals, and government and foundation funding? Her model assumes EA’s funding share decays from ~95% today to ~5% over thirty years as traditional science funders pick up the field, with stable annual budgets reaching ~$100M. The headline cost-effectiveness comes out to ~13 animal-years per dollar – within the same range as the 9–120 chicken-years per dollar Saulius Simcikas estimated for corporate cage-free campaigns. The argument I find persuasive in Graham’s piece isn’t the precise number; it’s the leverage. EA dollars spent on welfare-biology research now don’t just buy research; they pull in non-EA dollars later, the same way EA’s farmed-animal welfare investments eventually pulled in major-foundation and government attention.
Broad-Spectrum Existing Levers
The second tier is large-scale, high-confidence, well-understood existing donation targets. Their welfare effect on soil fauna might be enormously larger than their direct effect on the intended beneficiaries (depending mostly on nematode welfare ranges), but it runs through the same well-studied land-use lever, so we can be unusually confident in the direction of the effect.
Update: It’s important to note that Grilo has found a study that found surprisingly similar numbers of nematodes in the soil of different habitats. If true, this might ruin the robustness of this lever.
Funding GiveWell’s top charities. Grilo (2025a) estimates that GiveWell’s top charities increase cropland by ~137 m²-year per dollar, translating to 1.11 kQALY per dollar when accounting for soil animals – 1.74× the past cost-effectiveness of the Shrimp Welfare Project’s Humane Slaughter Initiative (HSI). The mechanism: saving lives increases the human population, which increases food demand, which increases cropland. Grilo estimates a 0.0157 human-year-per-dollar increase in human living time. But note that Roodman (2014) puts the sign of the mortality-fertility effect in some doubt.
Funding CEARCH’s High Impact Philanthropy Fund (HIPF). The Centre for Exploratory Altruism Research (CEARCH) estimates HIPF is ~12× as cost-effective as GiveWell’s top charities for human welfare. Grilo translates this into 13.4 kQALY per dollar with soil animals included – 20.9× HSI’s past cost-effectiveness. The same caveat as above applies, with the additional uncertainty Grilo flagged in his June 2025 update about whether HIPF’s chronic-disease-policy grants actually increase food consumption.
Chicken welfare campaigns. Broiler welfare and cage-free campaigns increase agricultural land by 93.9 and 16.4 m²-year per dollar respectively, because reformed systems require more feed per unit output. Grilo estimates these benefit soil animals 458× and 29× as much as they benefit chickens, though uncertainties over welfare ranges apply because most of them are nematodes. These chicken welfare reforms are likely good on their own terms; the soil-animal addition is a robust positive if soil animals have net negative lives and verly likely desirable from an asymmetric perspective.
Biofuel subsidies as a natural experiment. The WAWID shallow report on the US Renewable Fuel Standard (RFS) gives us a real-world confirmation of part 2’s mechanism: a policy-driven 2.1 Mha conversion of grassland to cropland produced an estimated reduction of ~12.9 trillion macroinvertebrates per year. The report’s own assessment is that movement counterfactual influence on the RFS is essentially zero – corn-state political coalitions and infrastructure lock-in mean nothing welfare-biology advocacy does will move that policy. So it isn’t an advocacy target. But it’s the cleanest empirical demonstration that the part-2 mechanism produces what part 2 says it produces, and worth knowing about for two reasons. First, EA-adjacent environmentalists who oppose the RFS on conservation grounds are working against welfare on this analysis; that’s an alignment worth noticing. Second, for asymmetric antifrustrationists worried about instrumental harm, the RFS structurally passes the second filter above: its costs to existing beings are bounded (food prices, displaced soy expansion), and no new sentient beings are created in service of the antifrustrationist gain. As such it can serve as a target for opportunistic altruism, in case a policy window does open.
Targeted Birth-Prevention
The most interesting addition from the WAWID is a small group of interventions that combine high per-individual suffering severity, plausible birth-prevention mechanisms, and clean structural-asymmetry profiles. None has the broad-spectrum coverage of the donations above; none affects both nematodes and arthropods. But they are the cleanest single-shot applications of the framework to specific species whose lives we have unusually strong reasons to think are net negative.
New World Screwworm continental suppression. The WAWID shallow report on suppressing Cochliomyia hominivorax across the Americas is, on my read, the single strongest entry in the database. C. hominivorax larvae cause myiasis by feeding on the living tissue of warm-blooded mammals – the Welfare Footprint Institute Pain-Track gives 10–30% excruciating pain and 70–80% disabling pain over the 14 days from oviposition to death by sepsis or toxemia. Mortality is near 100% in untreated wild infestations, and 1–4% of warm-blooded wild animals in endemic regions are infested at any given time.
The 1950s–2000s campaign eradicated NWS from North and Central America using sterile insect technique (SIT). The Panama biological barrier collapsed in 2024, the fly is spreading north again, and the United States is now scaling sterile-fly production toward 300 million releases per week to push it back to Panama. Continental suppression – proactively eliminating NWS across South America rather than perpetually defending a barrier – is feasible with new male-only CRISPR strains and gene drives, and the political momentum from the 2024 outbreak creates a 5–10 year window. Screwworm Free Future is the dedicated organization.
Why this is exceptional on the asymmetric antifrustrationist framework: the suppression intervention is doubly birth-preventing. It prevents future cohorts of NWS flies in the wild (the obvious channel), and it ends the perpetual sterile-fly rearing program – currently ~100M flies/week, scheduled to scale to 300M/week, and rising indefinitely under climate change. Almost no other intervention reduces both wild-population births and movement-driven captive-population creation. The Pareto profile is also unusually clean: warm-blooded wild animals win, livestock win, humans win, and the welfare cost to NWS flies themselves is bounded by RP’s welfare-range estimates – ~0.013 (using Black Soldier Fly as a proxy) versus mammalian ~0.5.
Gene drive for Anopheles gambiae. The WAWID shallow report on gene drives for malaria explores two architectures: a suppression drive that pushes the A. gambiae population toward zero, and a replacement drive that leaves population sizes intact but renders the mosquitoes unable to transmit Plasmodium. The shallow report leans toward replacement on extinction-ethics grounds. From my framework, suppression dominates: trillions of A. gambiae per year will continue to die of natural causes for as long as the species exists in malaria-endemic Africa. A successful suppression drive prevents the births of all those future cohorts, and the human side of the trade – ~600,000 fewer malaria deaths per year, mostly children – is enormous and aligned. The lever for a welfare-biology movement isn’t building the drive; it’s shaping the funding and regulatory criteria so welfare-considerate designs are favored.
Mating disruption for rice yellow stem borer and spongy moth Stop the Spread. Two structurally similar Lepidopteran interventions belong together. The WAWID shallow report on Scirpophaga incertulas describes pheromone-based mating disruption as a non-lethal alternative to insecticides in South and Southeast Asian rice paddies. Field trials in Indonesia produced 40–56% fewer insecticide applications and yields equal to or higher than insecticide controls. The shallow report estimates 6.7 × 10⁹ to 1.4 × 10¹² yellow stem borer larvae per year on the ~73.8 Mha of insecticide-managed rice in the region, plus a much larger uncounted reduction in non-target insect deaths from displaced organophosphate, pyrethroid, neonicotinoid, and diamide applications – all of which the shallow’s pain table classes as “likely excruciating,” with deaths over hours to days. Provivi and Syngenta announced commercial deployment in India and Indonesia starting in 2026; ASSIST’s “Phero Rice” project is already running through Farmer Field Schools in Vietnam.
The RP report on spongy moth outbreaks estimates 35–385 trillion Lymantria dispar dispar caterpillars suffered through outbreak years 2 and 3 in the United States over the last 50 years. Most die of starvation, Entomophaga maimaiga fungal infection, or LdNPV virus (“larval melting”) rather than from active control. The two intervention arms that align with asymmetric antifrustrationism are technologies that kill spongy moth eggs before hatch and mating disruption at the invasion front; both are pure birth-prevention. Faster Bacillus thuringiensis var. kurstaki (Btk) alternatives are death-improvement, which matters but ranks lower on the framework.
A note on vertebrate analogs. The WAWID shallows on a combined contraception/TB vaccine for New Zealand brushtail possums and on fertility control for white-tailed deer show the same logical structure on a smaller scale: birth-prevention via reproductive disruption, replacing a more painful counterfactual (1080 poisoning, hunting/culling) with a less painful one. Per-individual welfare ranges are higher for vertebrates, but population scales are bounded (~30M possums total, ~5M killed/year for 1080). Worth knowing about; not the largest priorities.
Personal Choices
Eat cheaply, donate the savings. Two arguments converge on the same recommendation, but they are independent and worth stating separately.
The cost-effectiveness argument is Grilo’s (2025b) own: a whole-food plant-based diet is cheaper than a standard one, so it allows him to donate more, and the donation impact dominates the dietary effect. On the per-kg numbers he computed:
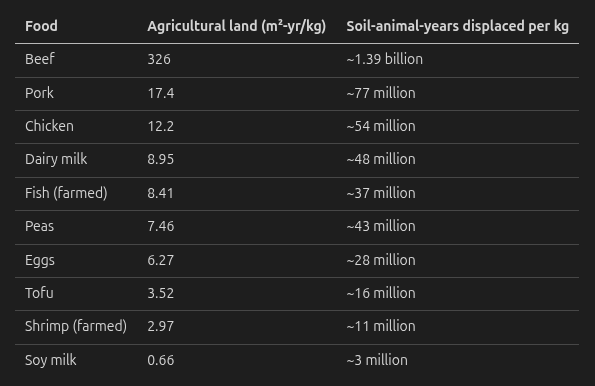
Beef looks like an outlier – ~26× more soil-animal-years displaced than chicken, ~395× soy milk. On the cost-effectiveness story alone, this is what Grilo means when he calls the dietary choice “ambiguous”: a beef-heavy diet displaces enormous amounts of soil-fauna life.
The soft Pareto argument is the other reason to eat plant-based, and it doesn’t depend on the sign of soil-fauna welfare. Eating beef creates cattle whose entire frustrated lives enter the welfare ledger at full weight, in order to displace soil-fauna lives via the cattle-feed → soy-expansion pathway. That’s exactly the cow → soy structure the second filter above, the soft Pareto filter, discourages: bringing new sentient beings into existence as the means of producing antifrustrationist gains. Plant-based calorie shifting between plant foods is a redirection of existing land use without creating new sentient beings to instrumentalize the gain. Eat-cheap-donate-the-savings works through the second mechanism only.
The two arguments converge on plant-based, but only the soft Pareto one distinguishes “create cow lives to prevent insect lives” from “redirect existing land use to prevent insect lives.” This is a place where the cost-effectiveness framing alone would lead you to cause greater instrumental harm – with the caveats mentioned above.
Robustness: Why Broad-Spectrum Approaches Win
A recurring theme: interventions that reduce both nematodes and arthropods simultaneously are more robust than ones that target one group, for three reasons.
Robustness across welfare ranges. The dominant uncertainty in Grilo’s model is the welfare range of nematodes. Rethink Priorities has estimated welfare ranges for some arthropod species, but nematode ranges have not been estimated. C. elegans has 302 neurons; soil mites and springtails have thousands; ants hundreds of thousands. Neurons are a crude proxy for welfare range, but it stands to reason that a nematode’s range may be much smaller than an arthropod’s. Conversely, though, nematodes are much more numerous. An intervention that affects both in the same direction is positive across this entire space.
Robustness against trophic backfire. Many soil-mite species are voracious nematode predators – some consume hundreds per day. If you selectively suppress mites, nematode populations can increase through predation release. Springtails compete with nematodes for fungal substrates. Broad-spectrum land-use change avoids these feedbacks because everything goes down together.
Robustness through existing policy levers. Land use, agricultural subsidy, and climate policy have massive existing political infrastructure. Building novel taxon-specific tools (nematode-only biocides, say) means starting from zero. Working through existing levers is enormously cheaper.
The targeted birth-prevention interventions are narrower on this axis than the broad-spectrum donations: they’re picked for severity of per-individual suffering, not breadth of taxon coverage. The portfolio reasoning says we want both kinds of bets.
What Not to Do
A handful of interventions look reasonable from a generic conservation or animal-welfare framing but turn out badly under asymmetric antifrustrationism.
Stabilizing oscillating biomes. Boom-bust ecosystems with predator-prey cycles, insect outbreaks, or fire-driven succession involve mass starvation and die-offs that look like prime suffering targets. The catch is that the available stabilization interventions tend to increase aggregate biomass and biodiversity – the stable state is typically more productive than the average of the cycle. From a conservation perspective that’s success; from an antifrustrationist perspective, more stable, more productive ecosystems support more potentially-suffering organisms, not fewer. Fire suppression is another example: preventing fire not only allows succession, which raises soil-fauna density and thereby aggregate suffering, it also stockpiles fuel for worse eventual fires. Either way, this is a bucket to exclude from the portfolio, not a bucket to fund.
Mandating wind-turbine curtailment for insects. The WAWID shallow report on climate-energy infrastructure estimates ~23,200 trillion insects per year killed by wind turbines in instantaneous, low-pain collision deaths. Population-dynamics arguments suggest most of this mortality is density-compensated – the population would have been the same without turbines. From the asymmetric antifrustrationist view this is one of the least problematic mortality sources on Earth: a near-instant death substituting for what would otherwise be parasitism, fungal infection, or starvation. The intervention proposed in the shallow report – mandating curtailment of turbines during peak insect activity – would effectively impose more painful counterfactual deaths to save individuals whose populations would not be reduced. Photovoltaic panel mortality is a different story: polarotactic insects are attracted to the panels for oviposition, and the eggs and larvae die on the surface over hours. It’s unclear how sentient they are at that stage, but the process doesn’t change the NPP of the area, so the population size will barely decrease. The white-grid panel design tweak that breaks the polarized-light trap is probably good. So: panel design good-ish, turbine curtailment bad.
Banning neonicotinoid seed treatments. The WAWID shallow report on reducing neonicotinoid seed treatments (NSTs) is unusually direct: the author concludes that the net welfare effect of NST bans “may be negative.” The reasoning lines up neatly with part 2’s pesticide framework. NSTs are systemic and persistent; they make plant NPP physiologically inaccessible to phytophagous insects, which is genuine carrying-capacity reduction rather than constant-NPP killing. Counterfactual deaths for the insects that would otherwise have eaten treated plants are dominated by parasitoids, which the shallow report describes as “possibly more severe and longer than death by acute NST exposure.” The shallow report even cites evidence that sublethal NSTs function as fertility control. NST bans typically replace NSTs with foliar pyrethroid or diamide sprays, both of which are worse on each of these dimensions. Yet conservation NGOs and several US states are converging on NST bans as a “win for invertebrates.” On this analysis, that convergence is wrong.
There’s a pattern across the three. Conservation-default and welfare-default intuitions diverge cleanly from asymmetric antifrustrationist conclusions, and the divergence isn’t accidental: conservation values aggregate populations and biodiversity, asymmetric antifrustrationism values reduced expected suffering, and these come apart whenever you can choose between “more healthier organisms” and “fewer organisms full stop.”
It doesn’t have to be that way. No one is just a “conservationist” and most people have empathy, especially people who go into ecology. It’s just a matter of sensitizing them to the fact that conservation and welfare are not or rarely correlated.
There is a framing around autonomy that seems internally consistent to me: Imagine a parkour athlete has a bad fall next to you. They’re still breathing but unconscious and clearly badly injured. Some of my friends would argue that they should not call an ambulance or interfere in any other way because that would impinge on the athlete’s autonomy, and being unconscious, they cannot consent. But I think this is a niche view, and to apply it to some species (nonhumans) but not others (humans) would be speciesist.
Summary of Recommendations
Ranked by the joint score on importance, neglectedness, tractability, asymmetry-fit, and robustness to nematode-welfare-range uncertainty:
Fund welfare-biology research. Soil-animal welfare research at WAI and RP, Eckerström Liedholm’s insecticide research agenda, and field-building infrastructure. Highest cost-effectiveness for me, robust to direction of effect, and the entire portfolio below depends on it. If you can fund only one bucket, this one.
Fund GiveWell top charities or CEARCH’s HIPF. Direct human benefit alone justifies the donation; the soil-animal benefit is a large addition if (probably when) the welfare sign comes out negative.
Targeted birth-prevention. New World Screwworm continental suppression (the surprise winner from the WAWID and probably the strongest single intervention available right now), gene drive A. gambiae suppression, mating disruption for rice yellow stem borer and spongy moth Stop the Spread. These are narrower than the broad-spectrum levers above but directly target species whose lives we have unusually strong reasons to think are net negative.
Don’t actively oppose biofuel mandates, wind turbines, or neonicotinoid seed treatments on conservation grounds. Each of these is doing antifrustrationist work as a side effect of mainstream policy. The cost in human welfare is bounded; the welfare benefit to wild invertebrates is enormous.
In part 4, I’ll take a detour into the philosophy of mind to ask what invertebrate suffering actually feels like – an answer that draws on the phenomenology of psychopathy and Buddhist insight practice, and that may reshape how we weight nematode versus arthropod welfare.

Hi Dawn. Thanks for the post.
---
I very much agree foundational research is the top priority. I also like the 3 concrete research directions you discuss, with the caveat that I would like field-building to focus much more on soil invertebrates.
---
"Converting natural biomes to agriculture reduces soil fauna density by 2–8×, and the sheer number of organisms affected (billions per m²-year) overwhelms everything else."
How did you get to "billions per m²-year"? Did you mean billions per $ donated to the interventions saving human lives the most cost-effectively? I estimated GiveWell's top charities decrease the living time of soil animals by 539 M animal-years per $ (https://forum.effectivealtruism.org/posts/WbmDhpqKcT8gjwpso/saving-human-lives-at-the-lowest-cost-increases-animal) (https://docs.google.com/spreadsheets/d/1r26jbZOSy6Cyojg8fPP-gGzk_pQzNKcIxemKNEsiVP0/edit?gid=631926438#gid=631926438&range=S1:S2), although I am very uncertain about whether they increase or decrease the living time of soil aninals (https://forum.effectivealtruism.org/posts/WbmDhpqKcT8gjwpso/saving-human-lives-cheaply-is-the-most-cost-effective-way-of?commentId=yx6pS5mbjksiGp8fg). The vast majority of soil animals are nematodes (https://forum.effectivealtruism.org/posts/ehrmin93mzseQMj82/total-number-of-neurons-and-welfare-of-animal-populations#Population1), and maybe these have a life expectancy at birth of around 20 days (https://www.abc.net.au/science/articles/2011/05/12/3214128.htm?site=tv&topic=latest). So I calculate GiveWell's top charities decrease the number of nematode lives by 9.84 billion per $ (= 539*10^6*365.25/20).
---
"The current cost-effectiveness numbers, including the welfare range of 6.68 × 10⁻⁶ for nematodes that Grilo deferred to Gemini 2.5 to estimate, are placeholders waiting for empirical replacement."
I am very uncertain about the expected welfare range of soil nematodes. For expected welfare range proportional to "individual number of neurons"^"exponent", and "exponent" from 0 to 2, which covers the best guesses that I consider reasonable, I estimate the expected welfare range of nematodes ranges from 7.79*10^-18 to 1 (https://forum.effectivealtruism.org/posts/ehrmin93mzseQMj82/total-number-of-neurons-and-welfare-of-animal-populations#Welfare_range1) (https://docs.google.com/spreadsheets/d/1S7ivkkC8jKRcWU3qj7PPTiEi4YiZDVbnnW1zCljscgE/edit?gid=1742797903#gid=1742797903&range=B1:P1).
Less importantly, Gemini's guess that a modal soil nematode has 240 neurons has a negligible impact on my estimate for the expected welfare range of nematodes you mention above (https://forum.effectivealtruism.org/posts/L9NZGB7xbxiwgndPk/welfare-biology-and-ai-the-quiz?commentId=wMx8KvXzPBdGMvc9M).
---
"The second tier is large-scale, high-confidence, well-understood existing donation targets. Their welfare effect on soil fauna might be enormously larger than their direct effect on the intended beneficiaries (depending mostly on nematode welfare ranges), but it runs through the same well-studied land-use lever, so we can be unusually confident in the direction of the effect."
I am confident that a greater increase in agricultural land per $ leads to larger changes in the welfare of soil animals per $. However, I have very little idea about whether increasing agricultural land increases or decreases the welfare of soil animals. I am very uncertain about whether increasing agricultural land increases or decreases the number of soil nematodes (https://forum.effectivealtruism.org/posts/WbmDhpqKcT8gjwpso/saving-human-lives-cheaply-is-the-most-cost-effective-way-of?commentId=yx6pS5mbjksiGp8fg), and I think effects on soil animals may be driven by effects on nematodes (https://forum.effectivealtruism.org/posts/WbmDhpqKcT8gjwpso/saving-human-lives-cheaply-is-the-most-cost-effective-way-of#Increase_in_the_welfare_of_soil_nematodes_as_a_fraction_of_the_increase_in_the_welfare_of_soil_ants__termites__springtails__mites__and_nematodes).
---
"Funding GiveWell’s top charities. Grilo (2025a) estimates that GiveWell’s top charities increase cropland by ~137 m²-year per dollar, translating to 1.11 kQALY per dollar when accounting for soil animals – 1.74× the past cost-effectiveness of the Shrimp Welfare Project’s Humane Slaughter Initiative (HSI)."
Here is my most recent analysis of the effects of GiveWell's top charities on soil animals (https://forum.effectivealtruism.org/posts/WbmDhpqKcT8gjwpso/saving-human-lives-cheaply-is-the-most-cost-effective-way-of). For expected welfare range proportional to "individual number of neurons"^"exponent", and "exponent" from 0 to 2, which covers the best guesses that I consider reasonable, I estimate a cost-effectiveness of 0.0123 to 135 M QALY/$ assuming theat increasing agricultural land increases the welfare of soil animals (https://docs.google.com/spreadsheets/d/1r26jbZOSy6Cyojg8fPP-gGzk_pQzNKcIxemKNEsiVP0/edit?gid=1730931468#gid=1730931468&range=EB1:EG1). As I said above, I have very little idea about whether this is the case or not.
---
"additional uncertainty Grilo flagged in his June 2025 update about whether HIPF’s chronic-disease-policy grants actually increase food consumption"
I estimated decreasing the consumption of sugar-sweetened beverages (SSBs) and sodium increases agricultural land 36.6 % and 96.2 % as much as it would if they decreased human mortality without any diet change (https://forum.effectivealtruism.org/posts/AL8xgPFhL2oD3MJsh/reduction-in-life-expectancy-and-agricultural-land-due-to).
---
"Grilo estimates these benefit soil animals 458× and 29× as much as they benefit chickens, though uncertainties over welfare ranges apply because most of them are nematodes."
I believe there is huge uncertainty in the welfare ranges of not only soil nematodes, but also in those of soil ants, termites, springtails, and mites (https://forum.effectivealtruism.org/posts/ehrmin93mzseQMj82/total-number-of-neurons-and-welfare-of-animal-populations#Welfare_range1).
---
"Biofuel subsidies as a natural experiment. [...] Second, for asymmetric antifrustrationists worried about instrumental harm, the RFS structurally passes the second filter above: its costs to existing beings are bounded (food prices, displaced soy expansion), and no new sentient beings are created in service of the antifrustrationist gain."
"The soft Pareto argument is the other reason to eat plant-based, and it doesn’t depend on the sign of soil-fauna welfare. Eating beef creates cattle whose entire frustrated lives enter the welfare ledger at full weight, in order to displace soil-fauna lives via the cattle-feed → soy-expansion pathway. That’s exactly the cow → soy structure the second filter above, the soft Pareto filter, discourages: bringing new sentient beings into existence as the means of producing antifrustrationist gains."
I think biofuel subsidies, and eating beef may increase the number of soil animals due to increasing the number of soil nematodes (https://forum.effectivealtruism.org/posts/pEbiEmeu2agEHJgyu/a-database-of-near-term-interventions-for-wild-animals?commentId=r7bCtdh8MWp2J9jqi).
---
"FoodAgricultural land (m²-yr/kg)Soil-animal-years displaced per kgBeef326~1.39 billionPork17.4~77 millionChicken12.2~54 millionDairy milk8.95~48 millionFish (farmed)8.41~37 millionPeas7.46~43 millionEggs6.27~28 millionTofu3.52~16 millionShrimp (farmed)2.97~11 millionSoy milk0.66~3 million"
This was supposed to be a table?